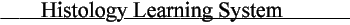
Common Confusions
The morphologies of certain tissues and organs can appear quite similar, making it difficult for a student to differentiate between them. The ability to successfully tell one tissue, or organ, cell, or organelle apart from another requires that you look for specific telltale features. Often those telltale features relate to the different functions of the two confusing structures. A basic approach to all identifications is to bear in mind the inseparability of:

Function is reflected in morphology and in location. As an example, consider the plasma cell. The plasma cell synthesizes and secretes antibodies: its function. Morphologically, it is characterized by a pale-staining cytocentrum where the ribosome-free Golgi apparatus packages those antibodies and an ample cytoplasm that is basophilic because of the presence of many protein-synthesizing ribosomes. Its ubiquitous location in connective tissues underlying epithelia is where antibody-producing cells are most effective in their role in the immune system.
Illustrated in this section are several commonly confused cells, tissues, and organs. The text in this section provides some functionally relevant suggestions on how to differentiate between and among commonly confused structures. It is good practice to use more than one functionally relevant feature to identify items: one for initial identification and one or two more to confirm that identification. The Taxonomic Key to Histological Organs (Appendix B) provides a more complete means for comparison among organs in tabular form.
A. Ultrastructure
 |
|
 |
|
 |
Smooth endoplasmic reticulum
(EM#2) |
|
Golgi apparatus
(EM#3) |
|
Rough endoplasmic reticulum
(EM#10) |
Each of these is a membranous intracellular organelle. The Golgi apparatus receives cell products (via vesicular transport), processes and packages them. Thus the Golgi apparatus consists of a series of flattened cisternae and associated vesicles that are typically organized with a distinct convex and concave side, reflecting its functional polarity. The smooth endoplasmic reticulum, despite its varied functions among different cells, is an anastomosing network of tubules with no obvious organizational pattern; the relative volume of SER varies with the cell's function. The membranous elements of rough endoplasmic reticulum are flattened cisterna (like pita bread) with surfaces studded with the ribosomes that produce the proteins destined for secretion (or for becoming integral membrane proteins).
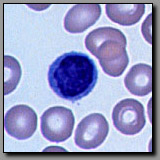
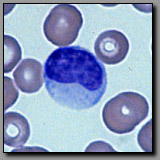
B. Cells
 |
|
 |
| Lymphocyte(#17) |
|
Monocyte(#17) |
Both of these cells occur in peripheral blood, and neither has a lobed nucleus. They both use the peripheral blood as a means of transport throughout the body. The small lymphocyte may be no more that 50% greater in diameter than the red blood cells; its nucleus is round and condensed and it has scant basophilic perinuclear cytoplasm. This morphology reflects its role in immunosurveillance, metabolically low-key until activated by an appropriate stimulus. The monocyte is nearly twice the diameter of a red blood cell; its nucleus is more lightly stained than the lymphocyte’s nucleus and may be indented; its ample basophilic cytoplasm often contains a few azurophilic granules (lysosomes). This cell is in transit from the bone marrow where it was born to the tissue where it will serve as a resident macrophage. Large lymphocytes, with more cytoplasm may be distinguished from macrophages by their more condensed nuclei.
 |
|
 |
|
 |
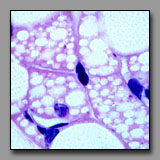
Multilocular (brown)
adipocytes(#49) |
|
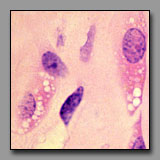
Macrophages
(#33) |
|
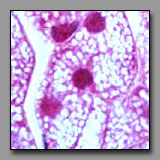
Steroid-producing cells
(#145) |
All of these cells contain multiple vacuoles of various sizes in their cytoplasm. In multilocular adipocytes and in steroid-producing cells these vacuoles are lipid droplets, and in macrophages they represent phagocytosed/digested material. In each of these cells, after standard tissue processing, the vacuoles can appear empty. These cells can be differentiated among themselves by location, cell shape, and cell packing. Macrophages often have an elongated irregular shape that reflects their ameboid, wandering nature and, although there may be many of them in one region, they are not contiguous. A macrophage displays its inclusions only as a result of substantial phagocytic activity, and so may not be easily recognized in the absence of such activity. Brown fat cells are compact in shape, assembled densely with others of their kind as part of a mass of tissue that has a high capillary density to enable effective transfer of heat from the cell’s metabolism to the blood, and eventually to the body. Steroid producing cells also assemble in cords or clumps where they have access to extensive capillary bed into which they deliver their hormones.
C. Tissues
 |
|
 |
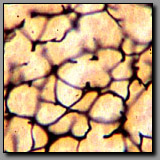
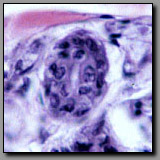
Reticular fibers-
silver stain(#70) |
|
Elastic fibers-
Verhoeff’s stain(#8) |
Special stains are required to specifically visualize reticular and elastic fibers. The commonly employed silver stain (for reticular fibers) and Verhoeff’s stain (for elastic fibers) both render the fibers black. The geometry of these fiber systems best differentiates between them. Reticular fibers provide a delicate scaffolding that supports densely packed cells, like those in a lymph node, liver, and spleen; they liberally branch and ramify around the cells they support. Elastic fibers provide recoil elasticity to the tissues in which they reside; they are relatively straight, often fine, and unbranched. Elastin, in the form of coarse wavy lamellae in the walls of blood vessels also stains black with Verhoeff’s stain.
 |
|
 |
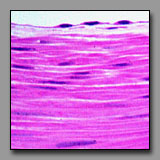
Stratified cuboidal
epithelium (#82) |
|
Transitional epithelium,
relaxed (#165) |
A stratified epithelium is classified according to the shape of the cells on its free or luminal surface. The surface cells of a stratified cuboidal epithelium are about as tall as they are wide, with appropriately circular nuclei. The surface cells of relaxed transitional epithelium are almost conical, with an apical surface that often bulges into the lumen; they are often considerably larger than underlying cells and may be binucleate. In this epithelium’s relaxed state the cells of the deeper layers appear squeezed and small. Five or more layers comprise this epithelium that lines much of the urinary system where the walls must adapt to very large changes in luminal volume. A stratified cuboidal epithelium, often found lining small ducts like those of sweat glands, typically has just two cell layers.
 |
|
 |
Stratified squamous
epithelium (#196) |
|
Transitional epithelium,
stretched (#166) |
Stratified squamous tissue gets its name from the shape of the cells comprising its surface (outermost) layer of cells: they are flattened. The cells beneath the surface layer are also relatively flattened, although the basal that which are arrayed along the undulating basal surface are often cuboidal. The presence of many cell layers in this tissue reflects its ability to protect underlying tissue from abrasive and wearing forces. When transitional epithelium is stretched the cells of its surface layer may also be flat like that of a stratified squamous epithelium. However there are two main differences between these two epithelia. With regard to the epithelial cells themselves, the stretched transitional epithelium has considerably fewer cell layers than the stratified squamous epithelium and the immediately underlying cells tend to be cuboidal rather than squamous. Also in the stretched transitional epithelium, the interface with the connective tissue below appears stretched and flat without the typical undulations that characterize the base of a stratified squamous epithelium.
 |
|
 |
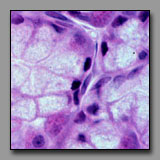
Endothelium
(#52) |
|
Mesothelium
(#166) |
Both of these are typical simple squamous epithelia, lining free surfaces. They differ in location. Endothelium lines blood vessels, and occurs on the concave or luminal side of tube-shaped vessels. Mesothelium covers the outside surface of organs that project into the body cavities and thus often covers the normally convex surface of an organ. Although both of these are simple squamous epithelia, the endothelial cells tend to have minimal cytoplasm such that there is little sign of cytoplasm between the bulging nuclei, while the mesothelial cells typically have more cytoplasm. It is noted that these two epithelia differ from other epithelia with regard to their origin from embryonic mesoderm.
 |
|
 |
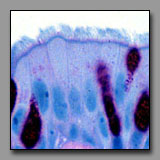
serous acinus
(#102) |
|
Mucous acinus
(#102) |
The cells of a serous acinus secrete proteins and glycoproteins, and their cytoplasm and inclusions reflect this. The nucleus typically appears rounded or oval and occupies the basal part of the cell (secretion granules are stored in the apical cytoplasm, from which they are released); the perinuclear cytoplasm often appears basophilic because of the presence of rough endoplasmic reticulum (responsible for protein synthesis). The cells of the mucous acinus synthesize and secrete mucinogen granules that contain an extensively glycosylated protein. This secretion product is water soluble and lost during routine tissue preparation so the cytoplasm of mucous cells appears empty in most H&E stained sections. The nucleus is usually flattened against the base of the cell by the voluminous stored product.
 |
|
 |
|
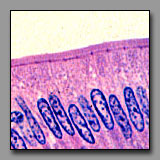 |
| Cilia (#131) |
|
Stereocilia (#169) |
|
Microvilli (#117) |
These luminal surface specializations have distinct functions: cilia are motile and move material in a direction parallel to the epithelial surface; microvilli increase a cell’s surface area and are absorptive; stereocilia also increase the luminal surface area and are absorptive (but in the inner ear, stereocilia serve as sensory transducers). The ultrastructural cores of microvilli and stereocilia are similar (bundled microfilaments), but in the light microscope, cilia and stereocilia appear to be the more similar structures. Both the cilia and stereocilia are individually (barely) discernable because there are fewer of them per cell than there are microvilli on the surface of an absorptive cell. With regard to relative height, the cilia are shorter (less than a quarter of the height of the cell) and more uniform in length than the stereocilia (which are more than half the height of the cell). Stereocilia tend to clump together distally like a paint brush, whereas the shorter cilia do not. Furthermore, as a reflection of the ultrastructural differences between cilia and stereocilia, a dark line of basal bodies appears in the apical cytoplasm beneath the cilia. The microvilli of absorptive cells are usually shorter (about 1 micrometer in length) than cilia and so tightly packed that they often form a solid-looking band (the striated border, or brush border) on the luminal surface of the absorptive epithelium.
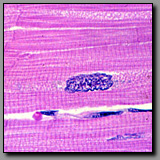 |
|
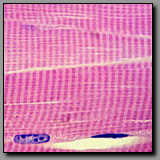 |
Cardiac muscle
(#63) |
|
Skeletal muscle
(#33) |
Both of these muscle types are striated, although the striations of cardiac muscle are less distinct because the cardiac myocyte cytoplasm contains many more mitochondria among its myofibrils. This high number of mitochondria coincide with the major metabolic demands of cardiac muscle. Cardiac myocytes are relatively small branching fibers, generally with one single nucleus per cell. Significantly, they are interconnected by intercalated discs that connect the individual cells electrically and structurally, enabling the cardiac muscle to behave like a large syncytium. Skeletal muscle fibers are long unbranching fibers with nuclei located peripherally so as not to interfere with the power and efficiency of contraction. The fiber size and multiple nuclei reflect the skeletal muscle fiber’s embryologic origin as a real syncytium of myoblasts.
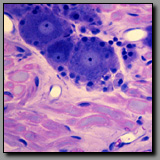 |
|
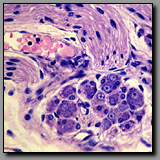 |
Dorsal root (spinal)
ganglion(#44) |
|
Parasympathetic ganglion
(#46) |
Both ganglia contain large neuronal cell bodies surrounded by nerve fibers; both are located in the peripheral nervous system. Neuronal cell nuclei are typically euchromatic and round-to-oval in shape and contain a distinct nucleolus. The cell bodies of the dorsal root ganglion (DRG) are larger than those of the parasympathetic ganglion because each of the former is metabolically supporting a vastly greater volume of axonal and neuronal cytoplasm than the latter. The ganglion cells of the dorsal root ganglion are of two sizes (large and small); they are unipolar and thus have smooth oval profiles. The parasympathetic ganglion neuronal cell bodies are all uniformly small; they are multipolar and thus more angular in outline, often with eccentric nuclei. Schwann cells encapsulate the neuronal cell bodies in both types of ganglia, but there are more satellite (Schwann) cells per DRG cell than parasympathetic ganglion cell because the surface area of the former is so much greater than that of the latter. As a reflection of differences in the extent of myelination that characterizes most fibers in the somatic system verses those of the visceral system, most axons associated with the dorsal root ganglion are heavily myelinated whereas those with the parasympathetic ganglion are unmyelinated or thinly myelinated. Finally, all dorsal root ganglia are located within intervertebral foramina (from which they are removed for histological processing), whereas parasympathetic ganglia are located in or near the organs they innervate.
 |
 |
 |
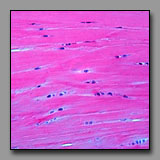 |
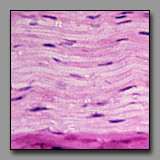
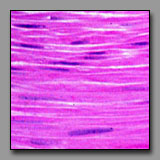
Peripheral nerve
(#43) |
Smooth muscle
(#52) |
Dense regular
connective tisse(#13) |
Fibrocartilage
(#32) |
All of these can appear to be linear, often undulating, eosinophilic tissues with dark nuclei interspersed. The peripheral nerve is typically most wavy in section because although not elastic, it must accommodate with increased length when the tissue in which it resides is stretched. Smooth muscle appears undulating only when it has been contracted (naturally or artifactually) to less than its initial length. The two connective tissues (dense and fibrocartilage) are usually too inflexible to appear undulating in tissue section. Peripheral nerve often has a foamy appearance because of the effects of routine fixation on the myelin (its lipid is usually not fixed, and is leached out of the section). The elongated nuclei in a peripheral nerve are those of Schwann cells situated peripheral to the myelin and of fibroblasts in the sparse endoneurium.
A key distinction between peripheral nerve and the others is that the peripheral nerve always has a cellular perineurium that controls the microenvironment within the peripheral nerve. Smooth muscle may also appear pale and wavy; its dark nuclei are distinctly intracellular; and there are rarely fibroblasts among them because smooth muscle synthesizes its own endomysium. The eosinophilic collagen of the dense regular connective tissue and the fibrocartilage is regular and densely packed. In tendons and ligaments, the nuclei interspersed among the collagen bundles are very sparse (more so than in the nerve or muscle) and they are all of fibroblasts (sometimes called tendonocytes). In fibrocartilage, the cells among the bundles of collagen are chondrocytes and they usually appear in linear clusters surrounded by some cartilage matrix. Unlike fibroblasts, the nuclei of the chondrocytes are more euchromatic and oval, and perinuclear cytoplasm is evident.
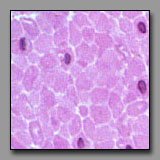 |
|
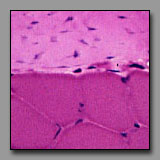 |
Smooth muscle
(#116) |
|
Dense regular connective tissue
and skeletal muscle(#43) |
In each of these transversely sectioned tissues, small dark nuclei appear against a field of eosinophilic material. The key to differentiating among them is to note the location of the nucleus with respect to the eosinophilic part. In the muscle tissues, extracellular space defines the limits of the cellular components. Transversely sectioned smooth muscle appears as a collection of circular eosinophilic profiles of varying diameters, representing transverse sections through different regions of the fusiform myocytes. The circular, heterochromatic smooth muscle nuclei are positioned within only the largest of these profiles (representing the middle of the myocytes). In transversely sectioned skeletal muscle, the nuclei are located at the periphery of the relatively large profiles of skeletal muscle fibers (whose striations are not usually apparent in transverse section). In dense regular connective tissue, the small heterochromatic nuclei are those of transversely sectioned fibroblasts and they are dispersed evenly throughout the packed eosinophilic field of extracellular collagen. There are also notable functionally significant differences in the relative density of the capillary beds of these three tissues: it should not be surprising that capillary profiles are very rare in the tendon, slightly more frequent in the smooth muscle, and quite numerous around each of the skeletal muscle fiber profiles.
D. Organs
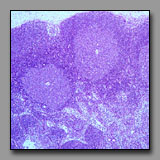 |
|
 |
|
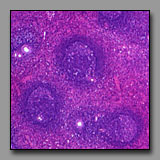 |
Lymph node
(#71) |
|
Thymus
(#74) |
|
Spleen
(#77) |
These three organs are encapsulated lymphoid organs and each is densely populated by lymphocytes. Each organ is characterized by the presence of noticeable morphologically different regions. In the case of the lymph node and the thymus, the two regions are a peripheral cortex and a central medulla; in the case of the spleen, the two somewhat intermixed regions are the white pulp (the part that is lymphoid tissue) and red pulp (densely populated by red blood cells). The lymph node and the thymus differ from each other notably in that the former has lymphoid nodules in the cortex, and distinct cellular cords and empty-appearing sinuses in the medulla; neither of these characterize the thymus. The cortex and medulla of the thymus differ principally in the relative packing density of the thymocytes. Hassall’s corpuscles in the medulla of the thymus are unique and diagnostic. The spleen, like the lymph node, has nodules with germinal centers, but there is no cortex and medulla. The splenic nodules are located within regions of white pulp in which may also be seen a central artery. The red pulp has cellular cords and sinuses like the lymph node, but because the spleen filters blood (and not lymph) splenic cords and sinuses may be filled with elements of blood (not lymph, as in the lymph node).
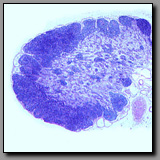 |
|
 |
Lymph node
(#71) |
|
Palatine tonsil
(#66) |
Both of these tissues contain lymphatic nodules. However, the lymph node has a distinct capsule on its convex surface and a cortex and medulla. The palatine tonsil is associated with a crypt, which is a branched invagination of the wall of the oral pharynx, lined with stratified squamous epithelium. On its concave surface, the tonsil is bound by an epithelium that is invaded by lymphocytes and on its deeper convex surface, a lamina propria of connective tissue. There is no medulla in the tonsil. Rather, this organ is simply lymphoid tissue intimately associated with the epithelium that it is monitoring.
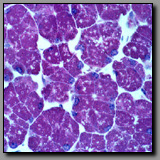 |
|
 |
|
 |
Parotid gland
(#101) |
|
Submandibular gland
(#102) |
|
Pancreas
(#104) |
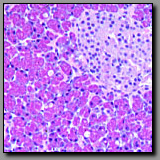
The parotid and submandibular glands are both solely exocrine glands, whereas the pancreas has both exocrine and endocrine components. The parotid and submandibular glands are major salivary glands with a similar pattern of ducts: intercalated, striated, interlobular, and excretory. These two glands differ in that the parotid gland has only serous acini, whereas the submandibular gland is mixed, having both serous and mucous acini, some of which are capped by serous demilunes. The exocrine portion of the pancreas, which as a major gland of the digestive system empties into the duodenum, is purely serous. Unlike the parotid gland there are no striated ducts in the pancreas; the duct system of the pancreas begins with a special portion that occupies the lumen of the acini. These intraacinar duct cells, called the centroacinar cells, secrete a bicarbonate-rich fluid that helps neutralize the chyme in the duodenum. The endocrine portion of the pancreas, the islets of Langerhans, are unique to the pancreas.
 |
|
 |
Pineal body
(epiphysis cerebri)(#144) |
|
Neurohypophysis
(posterior pituitary)(#140) |
Both of these endocrine glands are outgrowths of the brain and so it is not surprising that their histological appearance is similar. The similarity rests largely in the presence of a neuropil, a fine mesh of neuronal and neuroglial processes, and the presence of supporting cells that are unique to the central nervous system. Both of these organs, being endocrine glands, have prominent capillary networks. These glands differ in the location of their respective neurosecretory neurons: neurosecretory neurons are not present in the neurohypophysis. Axons that are engorged with neurosecretions (Herring bodies) synthesized in hypothalamic neurons are a distinguishing characteristic of the neurohypophysis. The principal cell type in the neurohypophysis is the pituicyte, the supporting cell. In the pineal, the neurons (pinealocytes) are the principal cells and they are clustered in cords or groups separated by supporting cells and neuropil. The unique and distinguishing characteristic of the pineal gland is brain sand, which unlike Herring bodies, is mineralized.
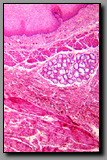 |
|
 |
|
 |
Esophagus
(#108) |
|
Vagina
(#196) |
|
Oral side
of lip (#96) |
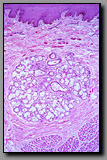
Each one of these has a prominent stratified squamous epithelium and an undulating interface with the underlying lamina propria. Distinguishing among them is based upon examining the whole tissue section. The esophagus is part of the tubular gastrointestinal system and it has the typical concentric layers of mucosa, submucosa, and muscularis externa. The vagina has the luminal epithelium and lamina propria in common with the esophagus, but none of the additional layers, as the vagina is a fibromuscular tube with indistinct inner circular and outer longitudinal muscle layers deep to the thick mucosa/submucosa. The lip, unlike the others, has some skeletal muscle (obicularis oris) deep to the mucosa, as well as some prominent mixed minor salivary glands. The side of the lip away from the mucosa has hair follicles and the keratinized epithelium of the face.
 |
|
 |
|
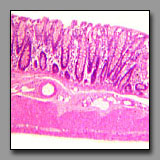 |
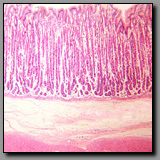
Fundic stomach
(#113) |
|
Pyloric stomach
(#114) |
|
Colon
(#112) |
These three regions of the digestive tract appear similar because of the numerous mucus secreting cells lining the lumen and the absence of villi. The geometry of the stomach mucosa differs from that of the colon in that the simple tubular glands in the stomach (gastric pits) have branched tubular glands emptying into them (gastric or pyloric glands), whereas the simple tubular glands of the colon (crypts of Leiberkühn) do not, and therefore occupy the full depth of the lamina propria. The lumen and gastric pits of the stomach are, in fact, exclusively lined with mucus-secreting cells, whereas the lumen and crypts of the colon have absorptive enterocytes scattered among the mucus-secreting goblet cells. The body and fundic part of the stomach can be distinguished from the other (cardiac and pyloric) regions of the stomach by the presence of the chief and parietal cells in the gastric glands. These two regions (body and fundic) comprise the major part of the stomach and their glands are the source of the enzymes and acid necessary for the stomach’s chemical digestion. The glands in both the cardiac and pyloric regions of the stomach (at the transition from the esophagus and to the duodenum, respectively) secrete mucus to aid in protecting the stomach lining from the acidic luminal contents. Cardiac glands are about the same length as the gastric pits into which they empty, whereas the pyloric glands are about twice the length of their gastric pits into which they empty.
 |
|
 |
|
 |
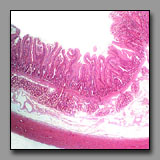
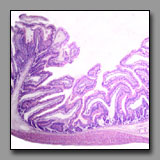
Duodenum
(#116) |
|
Jejunum
(#117) |
|
Ileum
(#120) |
These tissue sections all have intestinal villi. Intestinal villi occur only in the small intestine, the region of the digestive system where most of the absorption of nutrients occurs, and they increase the luminal surface area about eightfold. Although the geometry of the villi is not particularly easy to compare in histological sections, the villi progress from being tall, foliate, and numerous in the duodenum, to being less numerous and finger-like in form in the ileum. The ratio of goblet cells to enterocytes increases from duodenum to ileum. Both of these mucosal features correlate with maximum absorption taking place in the first part of the small intestine, and progressively less as the contents progress distally. A more obvious means to differentiate among these three regions lies in the submucosa: Brunner’s glands occur only in the duodenum (these glands are the major source of alkaline mucus to protect and lubricate the mucosa against the acidic gastric juices); and Peyer’s patches occur principally in the ileum (these clusters of lymphoid nodules in the lamina propria and submucosa monitor the luminal contents and mucosal surface); there is no such specialization in the jejunum, which is the middle portion of the small intestine.
 |
|
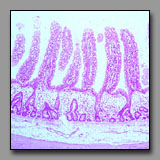 |
Gall bladder
(#156) |
|
Small intestine
(#120) |
These two organs may be easily confused because the simple columnar epithelium and what appear to be villi lining the lumen of the gall bladder appear to be of the jejunum (given that there are no Brunner’s glands or Peyer’s patches). To correctly differentiate between these two organs, examine the wall: the gall bladder has a simple mucosa/submucosa, no muscularis mucosae, a thin external muscular layer and, if not sharing a wall with the liver, a very thick subserosal layer. The gall bladder’s luminal epithelium consists of a single population of very tall thin columnar cells whose role is to concentrate the bile. What appear to be villi are mucosal folds that are particularly prominent in the nondistended state. The small intestine differs in several respects: it has regular villi and crypts; its columnar epithelium prominently displays both enterocytes and goblet cells; it has a distinct muscularis mucosae separating the mucosa from the submucosa; the muscularis externa has two distinct muscle layers, and the serosa is relatively closely applied to the peritoneal surface.
 |
|
 |
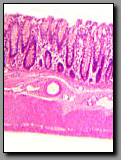
Colon
(#112) |
|
Appendix
(#124) |
There are no villi in these two sections of the gastrointestinal tract. The intestinal glands (crypts) are simple tubular and, because there are no branched glands emptying into them, these sections cannot be confused with the stomach. Two features enable distinction between these sections: only the colon has taenia coli, the prominent fiber bundles of the outer longitudinal layer of the muscularis externa (but best visualized in transverse sections), and only the appendix wall is dominated by lymphoid tissue that nearly obliterates the muscularis mucosae and reduces the number of crypts. Although discrete spots of GALT (gut-associated lymphatic tissue) typically occur in the colon, the GALT in the appendix infiltrates the entire mucosa and submucosa.
 |
|
 |
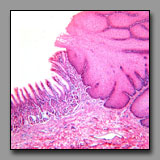
Esophagus/stomach
(#111) |
|
Anal canal
(#128) |
In these two regions of the digestive system there is an abrupt transition between stratified squamous epithelium and simple columnar epithelium. The most obvious difference between these two transition areas lies in the geometry of the section of mucosa with simple columnar cells. In the stomach, surface mucous cells form simple tubular gastric pits with smaller branched cardiac glands emptying into the pits’ bases. In the proximal portion of the anal canal, the epithelium of mostly mucous cells forms simple tubular glands (crypts). Differences are also apparent in the muscularis externa in this transition area: in the stomach the muscularis externa is robust and composed of smooth muscle; in the anal canal, the longitudinal layer of the muscularis externa becomes discontinuous whereas the inner circular layer forms the hypertrophied internal anal sphincter. Although there may be some skeletal muscle fibers in the muscularis externa of the esophagus (but only in the dog), skeletal muscle forms the large external anal sphincter that may be included in the anal canal tissue block, distal to the internal anal sphincter (smooth muscle). With respect to the stratified squamous epithelium: in the esophagus there is no keratinized portion, Whereas in the anal canal the nonkeratinized portion itself makes a transition to the keratinized epithelium of the skin beyond the anus. Finally, a serosa (visceral peritoneum) may be included in the esophagus/stomach section, but will never appear in the anal canal block.
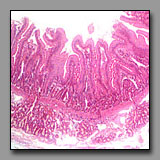 |
|
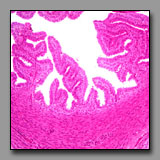 |
Duodenum
(#116) |
|
Ampulla of oviduct
(#186) |
These two sections may be confused because the villi of the duodenum are numerous and tall, making the apparent full thickness of the duodenal mucosa thicker than the wall’s muscle layers. Close inspection reveals the duodenum has all the standard components of the digestive system wall: muscularis mucosae, submucosa, muscularis externa with well-defined orthogonally oriented muscle layers, and a relatively thin serosa. The ampulla has a combined mucosa/submucosa, two indistinct layers of a muscularis, and a thick subserosal layer, which is derived from the mesenteric fold that supports the oviduct. The geometry of the duodenal mucosa consists of foliate villi projecting into the lumen and crypts invaginating the lamina propria, whereas the mucosa of the ampulla consists of longitudinal folds with secondary folds of mucosa. The diagnostic duodenal Brunner’s glands are not present in the ampulla.
 |
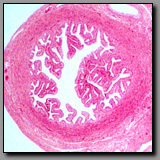 |
 |
 |
Ureter
(#163) |
Ampulla
(#186) |
Ductus deferens
(#173) |
Cervix
(#194) |
These four tubular profiles can be differentiated principally on the basis of their luminal epithelium. In each case the luminal epithelium reflects the unique function of the tube. The ureter is lined with transitional epithelium that characterizes most of the structures of the urinary system, reflecting its need to adapt to substantially changing luminal volumes. The ampulla of the oviduct has a simple columnar epithelium comprised of ciliated cells that beat to move the ovum or conceptus toward the uterus, plus secretory cells that provide material with nutritive and protective functions for the ovum. The ductus deferens, whose muscular wall is relatively the thickest wall of any tube in the body (to forcefully propel spermatozoa) is lined with a pseudostratified epithelium with long microvilli or stereocilia; the ductus deferens is a continuum of the epididymis where stereocilia serve to absorb some of the fluid initially necessary to transport the nonmotile spermatozoa. The cervix, a considerably larger structure than the other three, has a simple columnar lining epithelium that, like that lining uterine glands, consists of one cell type, a tall clear mucus-secreting cell. The cervix differs from the other three in that its wall is densely fibrous with relatively little smooth muscle (it propels nothing), and an outer epithelium, if present, is the stratified squamous epithelium of the vagina. The other three structures are ultimately covered with a mesothelium that encloses organs projecting into the peritoneal cavity.
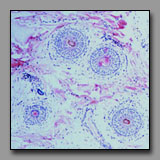 |
|
 |
Scalp
(#86) |
|
Ovary
(#148) |
Transversely sectioned hair follicles are multicellular structures that bear a superficial resemblance to multilaminar primary ovarian follicles because the stratified cells of the external root sheath look like cells of the membrana granulosa. Low power microscopic inspection is one means to differentiate between these two tissues. In the ovary, follicles are at different stages of development and so their morphology varies: primordial follicles with squamous follicle cells, unilaminar primary follicles with a single layer of granulosa cells, multilaminar primary follicles with multiple layers of granulosa cells, and secondary follicles with an antrum and cumulus oophorus. There is thus a range of ovarian follicle diameters and stages that is quite in contrast with a transverse section through hair follicles are relatively uniform in diameter and morphology, and uniformly dispersed in the section. Closer inspection confirms that the three-layered structure at the center of the hair follicle (hair) is not a single large oocyte. The scattered clumps of pale foamy cells seen in superficial sections through the scalp are sebaceous glands, and not to be confused with the foamy-looking discontinuous layer of theca interna cells encircling ovarian follicles.
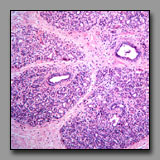 |
| 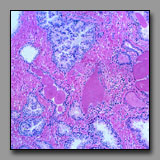 |
Mammary gland in
pregnant female(#90) |
|
Prostate gland
(#174) |
Both of these are compound glands embedded in a dense fibrous stroma. However, the mammary glands of a nonpregnant females are not active, and in fact, the growing alveoli are difficult to distinguish from the ducts. The adult prostate gland, in contrast, is actively secreting and this activity is reflected in the morphology of the tall pale secretory cells. At low power microscopic inspection, the distinct lobular organization of the mammary gland is evident, with branching interlobular and intralobular ducts between and within these lobules. The secretory elements of the tubuloalveolar prostate gland are separated by septa and lobules are indistinct. Lamellated prostatic concretions are unique to the prostate and, if present, are distinguishable from the homogenous eosinophilic product of the mammary gland.
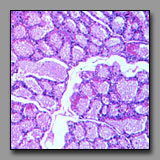 |
|
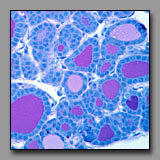 |
|
 |
Mammary gland-
lactating(#90) |
|
Thyroid
(#143) |
|
Prostate gland
(#176) |
These tissues may be confused because they each may have large follicles. They can be distinguished from one another primarily by examination of the shape and distribution of the follicles. The alveoli of the lactating mammary gland are in different stages of activity and those in one lobule or part of lobule may be drained by its lobular duct whereas others are engorged with product and not yet drained. Thus, in the mammary gland, the follicles of different sizes are clumped together. In the thyroid, the follicles of different diameters are intermixed (even disregarding the effects of tangential sections), and so a random distribution of sizes characterizes the thyroid gland. The secretory alveoli of the prostate gland may become so engorged with product, that they are commonly called follicles. However, the walls of the follicles are folded (to accommodate expansion) and so the follicular profiles are irregular and this is a feature not found in the thyroid or mammary gland. With regard to the stroma, the mammary glands are embedded in the breast tissue; there is no capsule and the connective tissue septa separating the lobules become quite narrow in the lactating gland. There is no duct system in the thyroid, an endocrine organ. The sparse interfollicular connective tissue accommodates a dense network of capillaries. In the tubuloalveolar prostrate gland, the stroma prominently surrounds the secretory follicles. Comparative microscopic examination of the secretory epithelia of these reveals that both the mammary gland and the prostate have typical secretory cells whereas the thyroid follicular cells are cuboidal to columnar cells that do not share the appearance of typical secretory cells because of the unique means by which its product is produced .
Go to:
[Front page||Main Index|Anatomical Figure|Schematic/Chart|Thumbnail|Slide Number|Keyword]
© 2002 Oxford University Press