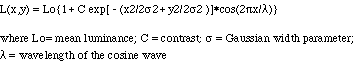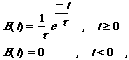| First- and second-order motion perception in Gabor micropattern stimuli: psychophysics and computational modelling Colin C. Clifford, Joshua N. Freedman, Lucia M. Vaina ContentsBack to First- and Second-order Motion INTRODUCTION Braddick [4] proposed that two processes, which he termed short- and long-range, underlie motion perception in humans. The short-range process was presumed to be mediated by simple motion selective filters with limited spatiotemporal support, while the long-range process was thought to operate by matching features over greater distances and longer time intervals. The use of the short-range / long-range distinction has been criticised on the grounds that the dichotomy might lie in the different types of stimuli that have been used to study motion perception rather than in distinct processing mechanisms [7,6]. Cavanagh and Mather [7] argued for an alternative classification which rests on the distinction between first- and second-order [8]. First-order motion is defined by spatiotemporal changes in luminance, second-order motion by stimulus attributes such as contrast, flicker and spatial frequency. However, much of the evidence supporting the existence of separate first- and second-order motion channels also involves a dichotomy in stimulus type, this time between stimuli in which motion is defined by changes in luminance and those in which motion is perceived in the absence of luminance cues. To demonstrate that the first-order / second-order dichotomy genuinely reflects the operation of distinct processing mechanisms, Boulton and Baker [2,3] devised a stimulus containing both first- and second-order information. Their stimulus consists of an array of Gabor micropatterns which are displaced between frames in a two-frame sequence, generating a percept of apparent motion. Boulton and Baker found that, for a dense stimulus array, direction judgements appear to be governed by first-order information, and for a sparse array by second-order. They also found that direction of motion can be reliably discriminated over only short ranges in the dense stimulus, but over much longer ranges in the sparse. It has subsequently been proposed by Lu and Sperling [13] that there are in fact three motion systems in human vision: a first-order system that uses a motion energy computation to extract motion from luminance modulations; a second-order system computing motion energy from texture and contrast modulations; and a third-order feature-tracking system. Lu and Sperling state that the nature of the motion computation in their third-order channel is as yet undetermined, but that motion energy analysis is a plausible candidate. We question the need to posit the existence of a dedicated second-order motion channel, in which motion extraction is preceded by texture grabbing, in addition to a feature tracking channel sensitive to motion defined by texture and contrast. As observed by Taub et al [16], many of the attributes which distinguish Lu and Sperlings second-order system from their third-order system are actually common to their first- and second-order systems. Rather than there being two short-range channels, extracting motion with and without a preceding full-wave rectification, Taub et al propose that there is a single short-range channel preceded by a more subtle non-linearity. We extend Boulton and Bakers study [3] to investigate the effects of using stimuli whose motion is defined by purely first-order or purely second-order information. Here, the first-order stimulus consists of luminance-defined Gaussian micropatterns rather than Gabor micropatterns. In the second-order stimulus, only the Gaussian contrast envelopes of the Gabor micropatterns are displaced, while their luminance-defined sinusoidal carriers remain static. We put forward a computational model which predicts psychophysical direction judgements. The model is a variant of those proposed by Chubb and Sperling [9] and Wilson et al [18], consisting of two distinct channels. Motion in both channels is extracted by a motion energy computation [1]. The filtering in the short-range first-order channel is linear up to the point of motion energy computation, so it is able to extract first-order motion but is blind to second-order. In the long-range second-order channel the stimulus is pre-processed by a low-pass texture grabber [10,17], rendering it sensitive to first- and second-order motion. The activities of the two channels are compared to establish which governs direction judgements for a given stimulus. In the rest of the paper we will use the terms first- and second-order channels in this way, referring to: a linear channel, operating at fine scales and sensitive only to first-order motion; a channel computing coarse scale motion energy subsequent to the action of a texture grabber. It should be noted that the second-order channel thus defined is different from the second-order sytem of Lu and Sperling [13] in that it analyses motion at a coarser scale than the first-order channel. PSYCHOPHYSICS Methods The following methods were adapted from Boulton and Baker [3]. The stimuli were generated on a Macintosh Centris 650 computer and presented on a Platinum Two Page Display SuperMac monitor which had a refresh rate of 75 Hz. A lookup table derived from photometer measurements was used to compensate for the nonlinear voltage-luminance characteristic. The stimulus raster size was 880x680 pixels (29.0x22.4cm), with 80 pixels subtending 1 degree of visual angle at the viewing distance of 150cm. The Macglib software package (Micro M-L, inc.) was used for developing the stimulus. The stimulus used in Conditions 1 and 3 was an array of Gabor micropatterns, each formed by the multiplication of a sinusoidal carrier and a two-dimensional Gaussian window:
The Gabor functions used had a mean luminance of 9.2 Cd/m2, a contrast of 0.20, a Gaussian width parameter of 0.33 degrees, and a wavelength of 0.44 degrees. These stimuli were presented in six rows, three above and three below a central fixation point. The individual micropatterns were positioned on a notional grid and jittered horizontally and vertically to prevent periodicity and clustering effects. The dense stimulus was made up of 11 micropatterns per row, giving a mean centre-to-centre spacing of 2.25
with parameters as for the Gabor micropatterns described above. COMPUTATIONAL MODELLING Methods We propose a two-channel model for the perception of first- and second-order motion. The first-order channel processes short-range motion information, while the second-order channel operates only at coarse resolution (long-range). The first-order channel analyses motion according to a simple first-order luminance-based scheme. In the second-order channel, the stimulus is first passed through a spatially and temporally low-pass filter and full-wave rectified. It is then analysed at coarse spatial resolution by a first-order mechanism identical to that used in the first-order channel. The first-order channel is insensitive to second-order motion, while the second-order channel responds to both first- and second-order stimuli. The architecture of the model is illustrated schematically in Figure 2. The initial stage of processing common to both channels is the removal of the mean luminance of the stimulus, which is assumed to occur at the retinal level in the human visual system [15]. This mean zero signal serves as the input to the short-range motion detectors in the first-order channel. The pre-processing stage in the second-order channel acts as a texture grabber [10], converting variations in the spatial and temporal frequencies of the image signal into variations in the magnitude of the pre-processed signal. Since the output of the second-order channel also varies with image contrast, it necessarily confounds spatial frequency and contrast by encoding their variations in a single response. Werkhoven et al. [17] provide psychophysical evidence that human perception of texture-defined motion is subject to the same confusion. Here we extend the spatial texture grabber proposed by Werkhoven et al. into the spatiotemporal domain by introducing a low-pass temporal filter. Motion is extracted by a standard first-order motion energy computation [1] using Gabor filters in space and time. Gabor filters were chosen to provide a measure of the amount of motion information available from the stimulus, not necessarily as an accurate model of the motion analysis filters in the human visual system. While there is evidence of spatial filters with receptive fields resembling Gabor functions in the primary visual cortex [13], the visual system's temporal filters will necessarily be causal and are likely to be better described by the modulation of an asymmetric temporal envelope than by a Gabor function [12]. Full details of the implementation of the model are given in the appendix. Results & Discussion Figure 3 shows the model's responses to sparse and dense two-frame apparent motion stimuli in each of the three stimulus conditions. The direction index of the response from each channel of the model is shown as a function of displacement. A direction index of +1 denotes motion in the direction of displacement; -1 denotes reverse motion; 0 indicates no net motion. Details of the calculation of the direction index are given in the Appendix. The layout of Figure 3 corresponds to that of Figure 1, with sparse and dense cases on the left and right and Conditions 1-3 going from top to bottom. Data points from the first-order channel are represented by circles, second-order by squares. To facilitate comparisons with the psychophysical data, solid lines are used to graph the response of the first-order channel to sparse stimuli and of the second-order channel to dense stimuli. For the sparse stimulus, results from the second-order channel in all three conditions are nearly identical: the direction index is close to 1 up to a displacement of 2.0 The results from the first-order channel differ greatly between the three sparse conditions. For Condition 1 (Figure 3A), the graph of the data from the first-order channel has a cyclical form with peaks at displacements of 0.75 For the dense stimulus, the results from both channels differ between the conditions. In Condition 1, the response of the first-order channel varies cyclically with peaks in direction index at displacements of 0.75-1.0 For all three conditions of the sparse stimulus, the response of the models second-order channel has qualitatively the same form as the psychophysical data (compare solid lines, Figure 3A-C with Figure 1A-C). The response of the first-order channel, however, bears no relation to the psychophysical data except in Condition 2, where it is very similar to the response of the second-order channel. We suggest that subjects perception of direction in the sparse stimuli is mediated by a second-order channel of the form implemented in the proposed model. For the dense stimulus, the situation is a little more complicated. Of the two channels, the response of the first-order (Figure 3D-F, solid line) more closely resembles the psychophysical data. However, there are two principal discrepancies. Firstly, the models first-order channel has an almost perfectly periodic response in Condition 1 (Figure 3D, solid line), with direction indices near +/-1 at all peaks and troughs, while the amplitude of the cyclical variations in the psychophysical data appears to increase with displacement (Figure 1D). Secondly, in Condition 3, the response of the first-order channel to small displacements ( < 1.25 Comparison of the psychophysical data and computational simulations suggests that perceived direction of motion in the sparse stimuli is mediated principally by a second-order motion mechanism, while for the dense stimuli both first- and second-order channels are involved. Why might the output of the second-order channel dominate for sparse stimuli but not for dense? We propose that the relative significance of the two channels' responses in determining the perceived direction of motion depends upon the relative magnitudes of the two channels' responses. In the model, we take as a metric of a channel's response its motion energy response summed over forward and reverse directions. This measure is independent of the direction index, as it depends upon the total motion energy regardless of direction. Direct comparisons cannot be made between the response magnitudes of the two channels as they are computed independently. The level of the response of the second-order channel depends upon the gain of the filters which pre-process the image as well as on the motion detection mechanism itself, and thus the absolute values of the responses of the first- and second-order channels cannot be compared. For this reason we look at the ratio of the response magnitudes of the first- and second-order channels (shown in Figure 4 for each of the six stimulus types). The values on the y-axis of Figure 4 may be scaled arbitrarily, corresponding to changing the gain of the texture grabber, without affecting the form of the graph. We see from Figure 4 that, for Conditions 1 and 3, the ratio of the response of the first- to the second-order channel is higher for the dense stimulus (filled squares and circles) than for the sparse (corresponding unfilled symbols). This is consistent with our suggestion that the perceived direction of motion in the dense stimuli reflects the responses of both channels, while motion perception in the sparse stimuli is governed by the second-order channel. The response ratios for both the dense and sparse stimuli in Condition 2 are lower than for the sparse stimulus in Conditions 1 and 3, suggesting that the response of the second-order channel determines perceived direction for both stimulus densities in Condition 2. However, the direction indices of the responses of the two channels to the stimuli in Condition 2 are very similar, so this latter point cannot be tested experimentally. CONCLUSION A two-frame motion stimulus was used to examine human perception of motion from first- and second-order cues. The psychophysical data obtained were modelled using a variant of the two-channel models proposed by Chubb and Sperling [9] and Wilson et al [18]. The concordance between our model simulations and the psychophysical data suggests that distinct mechanisms exist for the fine scale analysis of luminance-based motion and the coarse scale analysis of texture-based motion, with the relative activities of the two channels determining which mediates the perception of motion for a given stimulus. An interesting possibility proposed recently by Taub et al [16] is that the parallel motion processing pathways contain the same basic computational units and differ only in the scale at which they operate. If motion extraction is preceded at all scales by an asymmetric nonlinearity, then Taub et al speculate that the effective balance of the linear and nonlinear contributions might vary over scale, giving rise to apparent differences in the linearity of pre-processing between coarse and fine scales. A model with such an architecture has the attraction of parsimony, and cannot be discounted on the basis of our data. Acknowledgements This work was supported by NIH Grant EY-2ROI-0781-06 to LMV, and an NSF REU Grant to the Biomedical Engineering Department of Boston University for JNF. We thank Jane Boulton for providing the algorithm for the Boulton & Baker test, and Junzhen Lu and Jose Diaz for programming support. APPENDIX: IMPLEMENTATION OF MODEL AND SIMULATIONS The intial pre-processing stage common to both first- and second-order channels is the removal of the mean luminance, or DC component, of the stimulus. The mean is calculated over the entire image on a frame-by-frame basis. The signal in the first-order channel is then operated upon directly by the motion energy detectors. Texture grabbing Prior to the extraction of second-order motion, the image signal is pre-processed by a texture grabber. The operation of the texture grabber consists of three stages: initial linear filtering, full-wave rectification, and further linear filtering. Here, the first stage is low-pass filtering in space and time using a space-time separable filter. The spatial filter is a two-dimensional Gaussian, G(x,y):
The spatial extent of the filter is governed by the parameter, s, which here is set to 0.8 pixels. The two-dimensional Gaussian is itself separable into two one-dimensional Gaussians, so spatial filtering is carried out by successive convolution with two 3x1 masks. The spatial convolution wraps around at the edges of the image. The temporal filter, E(t), is a first-order low-pass filter with an impulse response which decays exponentially over time:
where t is the time constant. The value of t is necessarily positive, and gives a measure of the duration of temporal support. Here, t is set to 0.5 frames. For efficiency, temporal filtering is implemented recursively using the following recursion relation:
where T represents time sampled discretely in frames, I(T) is the input to the filter at time T, O(T) is the output of the filter, and b is a constant given by:
(For a derivation of the recursion relation see [11]). Motion energy computation
where sx, sy and st define the extent of the three-dimensional Gaussian envelope, kx and w are the spatial (horizontal) and temporal carrier frequencies, and fx and ft define the phase of the carrier relative to the envelope. Here, the preferred spatial and temporal frequencies, kx and w, of the filters are set to 0.1 pixels and 0.1 frames respectively, giving a carrier vertically oriented in space. The values of sx, sy are 2.5 pixels, and st is 2.5 frames, implemented in 15x1 convolution masks. The phases, fx and ft, are set to either 0 or p/2 to give the cosine and sine members of each quadrature pair. The outputs of these filters are then combined using trigonemetrical identities to give space-time oriented filters (see [1]).
where a is a constant given a small positive value to condition the quotient. The value of a is set at 10-7. If a was given a large value then this would qualitatively affect the form of D(x,y,t), as with low motion energies the directional response would be biased towards zero. However, in the simulations here the value of a was not manipulated as a parameter but fixed at a low value purely to prevent arithmetic exceptions in the execution of the model software.
Stimulus parameters
The Gabor functions have a carrier wavelength, lambda = 2pi/kx, of 8.0 pixels and the standard deviation, s, of the Gaussian envelope is 6.0 pixels. The centre-to-centre spacing of the Gabor patterns is 20.0 pixels in the dense condition, and 40.0 in the sparse. These values are in direct proportion to those used in the psychophysical experiments. The first image is present for 8 frames. The second image is simply a shifted version of the first, with wraparound at the edges, again presented for 8 frames. Jittering the positions of the micropatterns was found to affect the value of the direction index by a negligible amount (<0.05), so the simulations were run without jitter. Back to First- and Second-order Motion REFERENCES last update: 12/10/98 |